Table 1: Example for European types based on the numerical approach by a combination of three anthropometric traits (Lebzelter, 1929):
| Type | | | |
| Nordid | | | |
| Nordoid | | | |
| Atlanto Mediterranid | | | |
| Gracile Mediterranid | | | |
| Norid | | | |
| East Europid | | | |
| Dinarid | | | |
| Alpinid | | | |
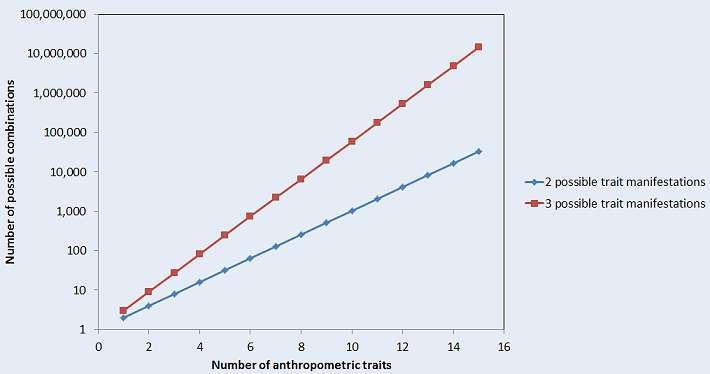
A cephalic index of at least 81 is considered short-headed, thus any tall, fair individual with a cephalic index of 80.9 would be classified as Nordid using this three-trait-based approach. With an index of 81 he would be Norid, ignoring other anthropometric features. The numerical approach was popular for example in the Polish and Yugoslav anthropology schools, which often used more than three metrics, but still only a limited number. Using numerical methods, sometimes Lappoid or South Mongoloid types were revealed among Native Americans or Australoid and Eskimoid types in Africa, even though these people have little relationship to Native Americans and Africans, respectively. This reveals a weak point of using an exclusively numerical approach: if only a handful of anthropometric traits is taken into account, huge differences in other features will not be considered and thus ignored. The numerical anthropologist is therefore highly dependent on his data and data quality. Also, with each metrical trait, the number of combinations quickly rises to huge numbers (see figure 1). Thus, a relatively small number of traits has to be selected. However, considering only 5 or 10 or even 50 anthropometric traits will not be sufficient to capture such a complex organism as a human, they can only capture a small fraction of a phenotype. Furthermore, measurement bias or different ways of measurement (e.g. there exist several methods to obtain the nasal index), or metrical traits altered by the environment will easily bias a classification. A human bias is also introduced easily by the choice of data acquired for the analysis. Thus, reasonable results with the numerical approach are only achieved with high data quality. Today, for instance, one could perform a 3D body scan and obtain thousands of anthropometric traits and add genetic info. This was not possible for classic anthropologists, and they didn't have the appropriate computers to handle such large amounts of data either. Either way, despite the drawbacks of the numerical approach, considering anthropometric features is helpful in any typological approach, one must only be cautious if no other information is considered.
Figure 1: Number of possible type combinations gives a specific number of anthropometric traits that have either two or three trait manifestations: